Hyaluronan - Protein Interactions
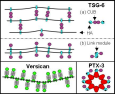 A cadre of proteins have been identified that interact with, crosslink, and bundle hyaluronan (HA) into unique structural and functional supermolecular complexes. These hyaluronan binding proteins, such as TSG-6, versican, and SHAP, have been thoroughly investigated in their interactions with short oligosacchardes of HA. Fewer studies have been performed to characterize their influence on longer HA molecules frequently present in tissues, or their effect on solutions of HA. Our research program aims to characterize the supermolecular aggregates and networks formed by protein-HA interactions using a variety of techniques, including microrheology, Fluorescence Recovery after Photobleaching (FRAP), and light scattering. This work is complemented by our studies of another common HA-protein structure in tissues, the pericellular coat.
A cadre of proteins have been identified that interact with, crosslink, and bundle hyaluronan (HA) into unique structural and functional supermolecular complexes. These hyaluronan binding proteins, such as TSG-6, versican, and SHAP, have been thoroughly investigated in their interactions with short oligosacchardes of HA. Fewer studies have been performed to characterize their influence on longer HA molecules frequently present in tissues, or their effect on solutions of HA. Our research program aims to characterize the supermolecular aggregates and networks formed by protein-HA interactions using a variety of techniques, including microrheology, Fluorescence Recovery after Photobleaching (FRAP), and light scattering. This work is complemented by our studies of another common HA-protein structure in tissues, the pericellular coat.
The domain of hyaluronan-rich biomaterials is largely uncharted territory. Biomaterials comprised of hyaluronan (HA) and/or its synthetic derivatives are widely used in tissue engineering and in many medical products including skin creams, arthritic treatments to repair synovial fluid in the joints and vertebrae, and ophthalmic surgeries for improved wound healing. HA is ubiquitous throughout body tissues and fluids. It is found in the extracellular matrix (ECM) and, attached to the surfaces of cells, in the form of the pericellular coat. In most cases, it associates with HA-binding proteins to form supramolecular aggregates and networks. Physiological studies relate HA-mediated biology to the structural and mechanical properties of these materials. Yet, no studies exist that map out the basic categories of HA-protein structures and their properties. A fundamental understanding of HA-protein biomaterials, including their structural organization and viscoelastic properties, will thus impact a multitude of biomedical applications as well as provide insights into fundamental biological processes ranging from cell adhesion and migration, inflammation, and ovulation.
Cables, networks and cell coats are among the reported HA-rich structures. Vital biomaterial properties are provided by many HA-protein structures. Very long HA-cables - crosslinked bundles of HA over 200 um in length, often appear in studies of inflamed tissues. It is now known that they are also made by a wide variety of cells as a response to ER stress. These cables have also been found to be pro-adhesive specifically for non-activated mononuclear leukocytes. Both TSG-6 and heavy chain proteins are suspected to crosslink these cables. However, concrete evidence of the crosslinking or its molecular origins does not exist.
Another example of crosslinked HA-aggregates appears in the synovial fluid, the biolubricant of joints. Synovial fluid's rheological properties are transformed when inflammation leads to the fragmentation and dilution of HA. A fascinating hypothesis is that the transient crosslinking of HA chains by proteins like TSG-6 and heavy chains might 'repair' the lost hydrodynamic properties of the larger HA molecules, providing a temporary fix to maintain the mechanical properties of the fluid.
The materials properties of the HA-rich cumulus matrix surrounding oocytes are vital for proper ovulation and fertilization. This extremely large, crosslinked HA network requires at least four HA-binding proteins for proper functioning, including TSG-6, heavy chain protein, and PTX-3. Its specialized viscoelastic properties purportedly protect the oocyte from mechanical squeezing during its release into the oviduct. The matrix is also believed to maintain the constant intrafollicular pressure needed for the extrusion. A rapid structural transition leading to the expansion of the cumulus matrix surface area for increased sperm attachment is associated with unregulated production of the HA-proteins.
The dearth of available data on HA-protein biomaterials is a reflection of the state of the field. Most HA-binding proteins have been identified in the past 20 years. Although great deal has since been uncovered about their molecular structures, little is known about the material properties of the diverse HA-networks they give rise to. This is in part because the typically small quantities of available protein are insufficient for most biophysical characterization techniques. Improved protein production capacity using recombinant molecular biology, the advent of adequate micro-volume physical characterization tools, and the numerous physiological observations of HA-protein structures make the time ripe for the studies proposed here.